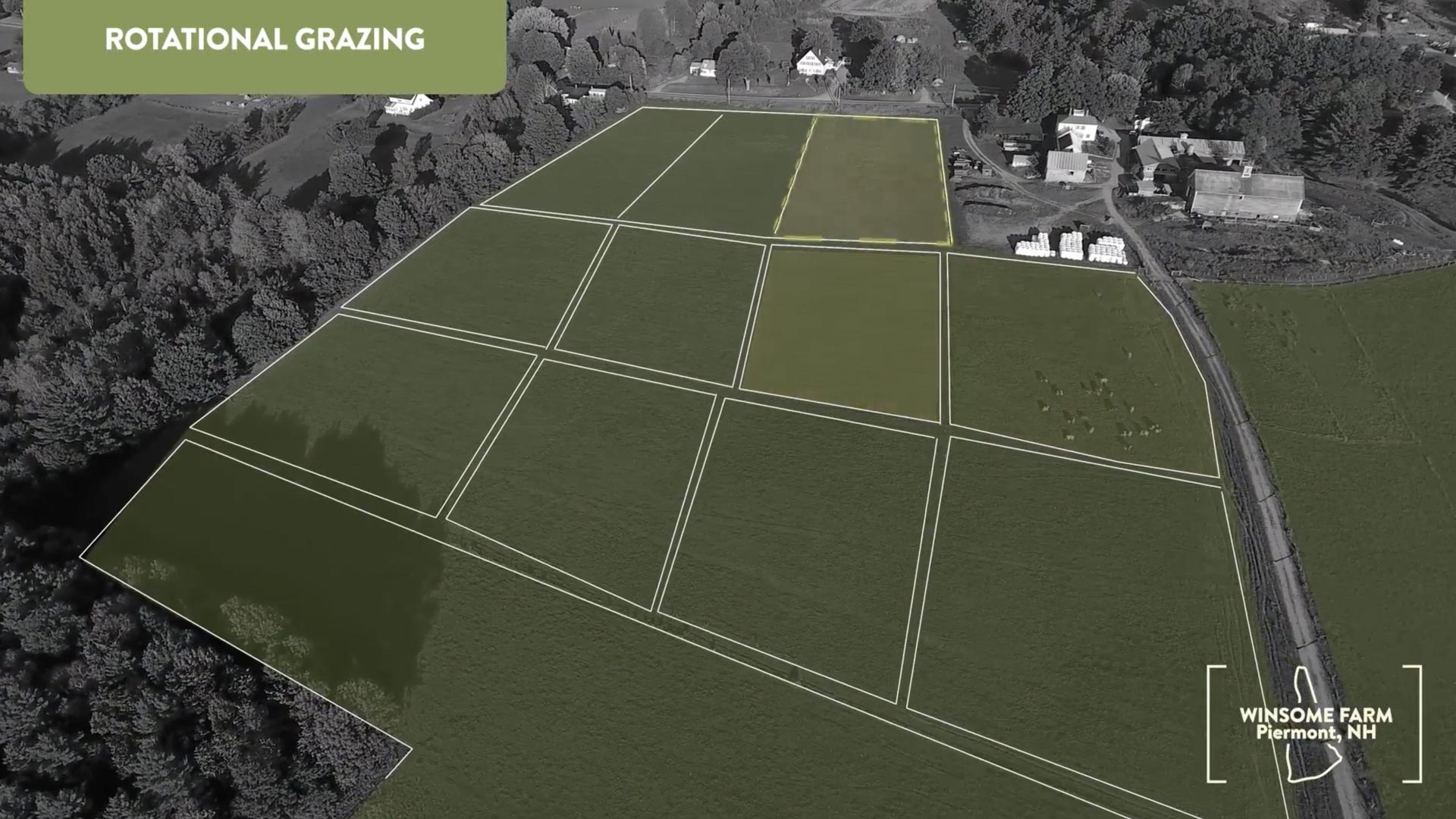
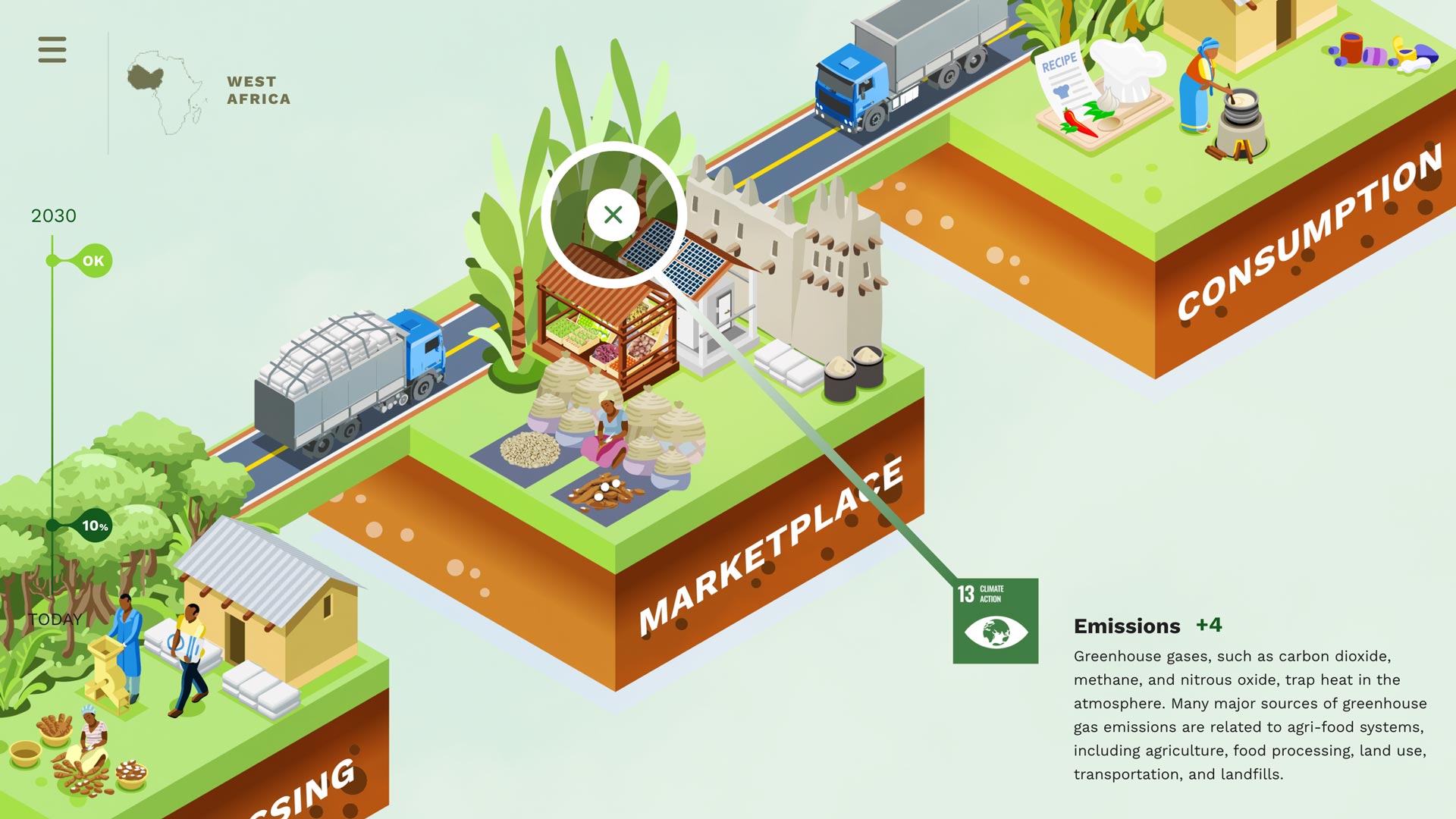
Can building an Ecological Benefits Framework for carbon markets accelerate our response to climate change?
An EBF (Ecological Benefits Framework) platform can untap the potential for blockchains to reward everyone across the value chain and standardize reporting practices for voluntary carbon markets.